Метаболизм ацетил-коэнзима а
Ацетил-КоАможет окисляться в цикле трикарбоновыхкислот. При этом обеспечивается 12 молекулАТФ на каждую молекулу ацетилкоэнзимаА. Ацетил-КоА может использоваться длясинтеза жирных кислот, при этом требуетсядостаточное количество энергии (АТФ).
При недостатке энергии, при нарушенииобмена углеводов и липидов синтезхолестерина через мевалоновую кислотунарушается и происходит образованиекетоновых тел с их накоплением в тканяхи выделением с мочой (кетозы). Мевалоновоякислотаявляется предшественником холестерина.
Однако реакции биосинтеза холестеринапочти полностью совпадают с реакциямибиосинтеза кетоновых тел. При нормальнойэнергетической обеспеченностиацетоацетил-КоА полностью превращаетсяв печени в мевалоновую кислоту, затемв холестерин.
При уменьшении углеводов,сахарном диабете, нарушении обменалипидов происходит увеличение кетоновыхтел, развиваются кетозы.
Пути образования кетоновых тел
1.Образование кетоновых тел происходитпри углеводном голодании, уменьшенииэнергетических источников для синтезажира, холестерина из активированнойуксусной кислоты. При этом активированнаямевалоновая кислота распадается наацетоуксусную кислоту и ацетил-КоА:
Ацетоуксуснаякислота частично восстанавливается до(β-оксимасляной кислоты, а частичнодекарбоксилируется, образуя ацетон:
II.Если CH3-CO-KoAобразуется много и недостаточно энергиидля синтеза жира, то образуетсяактивированная ацетоуксусная кислота:
Затем образуетсясвободная ацетоуксусная кислота:
Образующаясяацетоуксусная кислота может превращатьсяв β-оксимасляную кислоту или ацетон,как указано выше. Кетоновые теласинтезируются в печени и поступают вкровь. В нормальных условиях β-оксимаслянаякислота, ацетоуксусная кислота окисляютсяв тканях мышц и являются источникомэнергии.
Однако при нарушениях обмена(сахарный диабет), когда использованиеуглеводов затруднено и усиливаетсяокисление липидов, при длительномнедостатке углеводов, происходитчрезмерный биосинтез кетоновых тел собразованием ацетона.
При этом происходиткетонемия (ацидоз) и нарушение нормальногофункционирования организма.
Окислениененасыщенных жирных кислот происходитпосле их превращения в соответствующуюнасыщенную жирную кислоту. Затем ониокисляются по β-углеродному атому.
Окислениежирных кислот с нечетным числом углеродныхатомов происходит так же, как и приокислении жирных кислот с четным числоматомов углерода. Отличие состоит в том,что в конечном итоге образуетсяпропионил-КоА, который вначале подвергаетсякарбоксилированию, в результатеобразуется активированная янтарнаякислота (сукцинил-КоА). Сукцинил-КоАокисляется в цикле трикарбоновых кислот.
Биосинтез липидов
Способностьживотных запасать полисахариды довольноограничена; поэтому глюкоза, поступающаяв организм в значительных количествах,используется для синтеза жирных кислоти глицерина. Важным процессом являетсятакже биосинтез холестерина, так какон является исходным продуктом дляобразования многочисленных биологическиактивных стероидов.
Синтезвысших жирных кислот происходитиз ацетил-KoAв цитозоле. Фермент ацетил-КоА-карбоксилаза,содержащий биотин, катализируетАТФ-зависимый синтез малонил-КоА изацетил-КоА и СО2:
Приучастии ацилпереносящего белка (АПБ)Мм 10 Да происходит синтез ацетил-SАПБи малонил-SАПБ:
На следующем этапе ацетил-SАПБ и малонил-SАПБвзаимодействуют с образованиемацето-ацетил- SAПБ:
ацетоацетил-SАПБ
Послеэтого ацетоацетил-SАПБвосстанавливается по β-углеродномуатому с участием НАДФН2,в результате образуется β-гидроксибутирил-SAПБ.Затем происходит присоединениеактивированной уксусной кислоты имолонила к ацетилпереносящему белку(АПБ):
β-Гидроксибутирил~SАПБпод действием фермента β-гидрокси-АПБ-дегидратазытеряет молекулу воды (5) и превращаетсяв кротонил-SАПБ;последний с участием НАДФН2восстанавливаетсяв бутирил-SAПБ.Далее цикл реакции повторяется.
Еслиидет синтез пальмитиновой кислоты(C16),то кбутирил-SAПБприсоединяется последовательно еще 7раз малонил~SАПБ,при этом отщепляется дистальнаякарбонильная группа малонил~SАПБв виде CO2.Суммарный результат этих реакцийследующий:
ацетил-КоА+ 7 малонил-КоА + 14 НАДФН2
пальмитиноваякислота + 7CO2+ 8 HSKoA+ 14 НАДФ + 7 H2O
В этомпроцессе 7 молекул CO2образуется за счет свободных карбоксильныхгрупп 7 молекул малонил-КоА. Из 16 углеродныхатомов пальмитиновой кислоты 2 атомаобразуется за счет ацетил-КоА, а остальныеза счет малонил-КоА.
В ходесинтеза 7 карбоксильных групп – С=О-восстанавливаются из семи групп -CH2-, на это расходуется 14 НАДФН2:за счет 7 из них образуются водородныеатомы групп -CH2-, а за счет остальных семи – кислородкарбонильных групп превращается в воду.
Биосинтезфосфатидных кислот и триглицеридовпроисходит в печени и жировой ткани приналичии глицерина и жирных кислот.Глицерин образуется при гидролизелипидов, затем под влиянием ферментаглицерокиназы с участием АТФ синтезируетсяглицерофосфат:
Глицерофосфатможет образоваться также издиоксиацетонфосфата, возникшего приокислении глюкозы путем восстановленияНАДН2.
фосфодиоксиацетон глицерофосфат
Глицерофосфат далее используется для биосинтезафосфатидной кислоты. Реакция катализируетсяфосфатидсинтетазой:
глицерофосфат ацил-КоА фосфатидная кислота
Вфосфатидной кислоте в двух позицияхспиртовые группы глицерина соединенысложноэфирной связью с остатками жирнойкислоты (чаще всего с 16 и 18 углероднымиатомами), а в одной позиции с фосфорнойкислотой.
Синтез триацилглицериновосуществляется из фосфатидной кислоты.Вначале из фосфатидной кислоты отделяетсяфосфорная кислота, при этом образуетсядиацилглицерин.
Далее диацилглицеринреагирует с третьей молекулойактивированной жирной кислоты (ацил-КоА)и образуется триацилглицерин:
фосфатидная диацилглицерин ацил-КоА триацилглицерин
кислота
Биосинтез фосфоглицеридов происходит в эндо-плазматических мембранах клетки.Фосфатидилэтаноламин синтезируется вследующей последовательности: в началеэтаноламин фосфорилируется с участиемАТФ и фермента этаноламинкиназы:
этаноламин фосфоэтаноламин
Затем фосфоэтаноламин реагирует с ЦТФ, в результате образуется цитидилдифосфатэтаноламини пирофосфат:
фосфоэтаноламин+ ЦТФ → ЦДФ-этаноламин.
ДалееЦДФ-этаноламин взаимодействуя с 1,2-диацилглице-ридом, превращается вфосфатидилэтаноламин:
ЦДФ-этаноламин + 1,2-диацилглицерид →
→ фосфатидилэтаноламин + ЦМФ.
Биосинтезфосфатидилхолина (лецитина) осуществляетсяследующим образом: синтезируетсяфосфатидилхолин из фосфатидилэтаноламинапутем метилирования или же вначалехолин взаимодействует с ЦТФ.
ЗатемЦДФ-холин с 1,2- диацилглицерином образуетдифосфатидилхолин:.
ЦДФ-холин + 1,2 – диацилглицерин →
→ фосфатидилхолин + ЦМФ
Биосинтезфосфатидилсерина происходит путемобмена этаноламина на серин:
Фосфатидилэтаноламин + L-серин →
→ фосфатидилсерин + этаноламин.
Источник: https://StudFiles.net/preview/6050956/page:58/
Вещества природного происхождения, построенные из фрагментов молекул уксусной кислоты
Е.А.Кэри
Образование углеводов из воды и углекислого газа называется фотосинтезом и происходит за счет энергии Солнца, которая как бы «запасается впрок» в данном веществе. Фотосинтез протекает в присутствии зеленого пигмента растений – хлорофилла – и выражается уравнением:
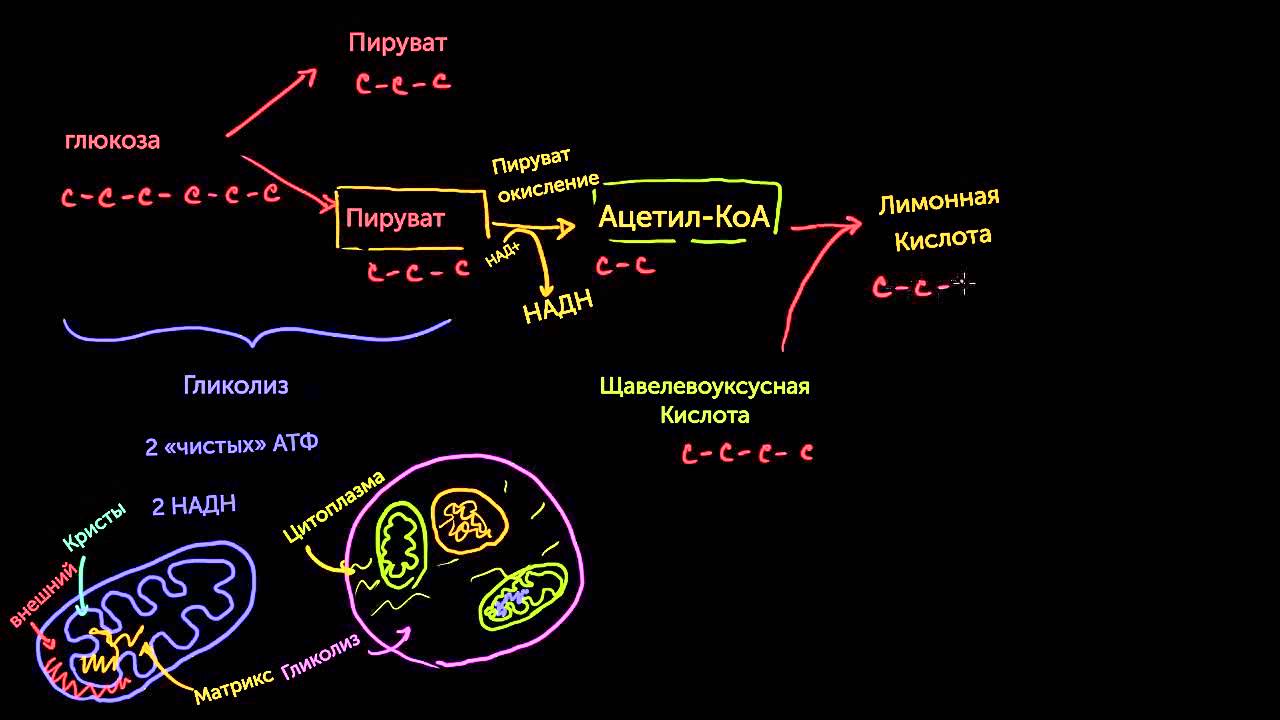
Важнейший углевод – глюкоза – используется клетками живого организма как источник энергии. Превращение глюкозы в пировиноградную кислоту, а затем в молочную кислоту называется гликолизом и сопровождается выделением значительного количества энергии:
Большинство биохимических реакций протекает при рН ~7. При таких рН карбоновые кислоты в основном находятся в форме солей (анионы RCOO–), а не собственно кислот RCOOH. Говоря точнее, гликолиз глюкозы происходит через промежуточное образование пирувата СН3С(О)СОО– и приводит к лактату СН3СН(ОН)СОО–.
Одним направлением метаболизма пирувата в живом организме является превращение его в более простые вещества (например, вода, углекислый газ) с одновременным выделением или образованием запаса энергии; другим – превращение в ацетильное производное с целью использования в качестве исходного вещества при биосинтезе более сложных соединений. Такие природные продукты, составленные из ацетильных фрагментов, играют ключевую роль в химии жизни.
Ацетильный коэнзим А
Ацетильная группа участвует в большинстве важнейших биохимических реакций в форме ацетилкоэнзима А.
https://www.youtube.com/watch?v=jyt3oimqzm8
Ацетилкоэнзим А является тиоэфиром. Его образование из пирувата протекает в несколько стадий, что можно записать общим уравнением:
Боковая цепь коэнзима А оканчивается сульфгидрильной группой –SН. Ацетилированием тиольного коэнзима А по группе –SН получаем ацетилкоэнзим А. Соединения типа RС(O)SR’ более подходящие переносчики ацильной группы чем кислородные аналоги RС(O)ОR’ (связь ацил-сера менее прочная).
Кроме того, у тиоэфиров RС(O)SR’ выше содержание енола в равновесной смеси эфир L енол по сравнению с эфирами RС(O)ОR’. Каждое из этих свойств наблюдается у ацетилкоэнзима А.
В одних случаях он действует как агент-переносчик ацетильной группы, в других выступает как нуклеофильный субстрат, тогда реакционным центром служит a-углеродный атом ацетильной группы:
Жиры и жирные кислоты
К веществам, синтезируемым в организме из структурных ацетатных единиц, относятся жиры. Они выполняют разнообразные функции в живых существах, в том числе являются источником энергии.
Если углеводы быстро восполняют потребности организма в энергии, то жиры «сгорают» сравнительно медленно. Зато жиры освобождают в два раза больше энергии, чем углеводы такой же массы. Организму выгодно запасать и сохранять энергию в форме жиров.
При этом меньшей массой вещества обеспечивается получение большей энергии.
Жиры представляют собой природные смеси триацилглицеринов или, что то же, глицеридов жирных кислот. Ацильные группы1 в триацилглицерине могут быть одинаковые или разные. Например:
В результате гидролиза жиров получают длинноцепочечные жирные кислоты и глицерин. Большинство жирных кислот природного происхождения содержат 12–20 атомов углерода в виде неразветвленной углеродной цепи.
Жирные кислоты входят в состав жиров растений и животных, где они образуются биосинтезом из ацетатных звеньев при участии ацетилкоэнзима А.
Биосинтез жирных кислот
Особенности биосинтеза жирных кислот хорошо видны на примере образования бутановой кислоты из двух молекул ацетилкоэнзима А. Проводником этого превращения служит комплекс энзимов, известный как синтетаза жирных кислот.
Часть комплекса, называемая ацильный переносчик белка (АПБ), содержит боковую цепь, похожую по строению на коэнзим А.
Важной начальной стадией биосинтеза жирной кислоты является перенос ацетильной группы от молекулы ацетилкоэнзима А к сульфгидрильной группе –SH ацильного переносчика белка:
Другая молекула ацетилкоэнзима А реагирует с диоксидом углерода (в действительности с гидрокарбонат-ионом при рН~7), образуя малонилкоэнзим А2:
Затем путем нуклеофильного ацильного замещения (вместо –SKoA становится –SАПБ) происходит перенос малонильной группы к ацильному переносчику белка:
После того как структурный блок, содержащий три атома углерода, присоединяется к АПБ, происходит образование углерод-углеродной связи между a-углеродом малонильной группы и карбонильным углеродом ацетильной группы
S-ацетилацильного переносчика белка. Возникновение новой связи С–С сопровождается декарбоксилированием и приводит к четырехуглеродному фрагменту ацетоацетильной группы, связанной с АПБ.
Этот процесс представлен ниже как стадия 1.
Затем ацетоацетильная группа превращается в бутаноильную группу в результате последовательных реакций, что изображено стадиями 2–4.
Стадия 1. Ацетильная группа переходит к a-углеродному атому малонильной группы с выделением СО2:
Стадия 2. Кетонный карбонил ацетоацетильной группы восстанавливается до спиртовой группы. Такое восстановление протекает при участии фосфатного эфира – никотинамидадениндинуклеотидфосфата (НАДФН):
Стадия 3. Дегидратация b-гидроксиацильной группы:
Стадия 4. Восстановление двойной связи a,b-ненасыщенной ацильной группы. Эта стадия требует присутствия коэнзима НАДФН:
Четыре углеродных атома бутаноильной группы перешли от двух молекул ацетилкоэнзима А; гидрокарбонат-ион, участвующий в реакции, не входит в состав конечного продукта (молекула СО2, что используется для превращения ацетилкоэнзима А в малонилкоэнзим А, регенерируется на стадии 1 при декарбоксилировании, сопровождающемся образованием углерод-углеродной связи).
Многократное повторение четырехстадийного цикла, приведенного выше, дает неразветвленные ацильные группы с 6, 8, 10, 12, 14 и 16 атомами углерода. В каждом случае удлинение цепи происходит за счет реакции с малонильной группой, соединенной с ацильным переносчиком белка.
Так, биосинтез 16-углеродной ацильной группы гексадекановой (пальмитиновой) кислоты записывается уравнением:
Наращивание в результате биосинтеза углеродной цепи жирной кислоты прерывается в случае переноса ацильной группы от ацильного переносчика белка к коэнзиму А. Образующийся ацилкоэнзим А готов участвовать в новых биологических превращениях, одним из которых может быть:
а) удлинение цепи, приводящее к образованию ацильной группы с более чем 16 атомами углерода; б) введение одной или нескольких двойных связей С=С;
в) ацильный перенос от серы к кислороду с образованием эфиров, например триацилглицеринов. Процесс, в котором молекулы ацилкоэнзима А превращаются в триацилглицерины, происходит с участием промежуточных соединений – фосфолипидов.
Фосфолипиды
Триацилглицерины (классические жиры) получают не путем ацильного переноса непосредственно к глицерину, а в результате последовательных превращений, первым среди которых является ацильный перенос к L-глицеро-3-фосфату.
Продукт реакции переноса двух ацильных групп к L-глицеро-3-фосфату называется фосфатидиловой кислотой:
В результате гидролиза фосфатидной кислоты по фосфоэфирной связи получают диацилглицерин, который, взаимодействуя с третьей молекулой ацилкоэнзима А, превращается в триацилглицерин:
Фосфатидная кислота является не только интермедиатом (промежуточное соединение) в биосинтезе триацилглицеринов, но и биопредшественником других представителей класса соединений, называемых глицерофосфатами. Фосфорсодержащие производные жиров, к которым относятся и глицерофосфаты, принадлежат к одному типу соединений – фосфолипидам.
Важнейшим фосфолипидом является фосфатидилхолин, называемый также лецитин:
Это смесь диэфиров фосфорной кислоты. Одна эфирная функция образована диацилглицерином, другая – холином [–ОCH2CH2N+(CH3)3].
Сокр. пер. с англ. А.Д.Микитюка
(Organic Chemistry. N. Y.: МсGraw-Нill Companies, Inc., 1996, 1151 p.)
________________________________________________________________________
1Ацильной группой называется группа атомов где R – углеводородный радикал. Этот радикал может быть насыщенным, ненасыщенным, ароматическим, содержать заместители, в том числе и гетероатомы Cl, O, S и т. д.
2Малонил
– ацильный остаток малоновой кислоты HOOCCH2COOH.
Источник: http://him.1september.ru/article.php?ID=200102201